Introducción
Una de las enfermedades parasitarias más importantes del ganado en todo el mundo es la
babesiosis bovina, enfermedad producida por protozoarios del género Babesia.
1 En México, 75% de las 23,316,000 cabezas de ganado vacuno
2 están en riesgo de adquirir babesiosis.
3
En este sentido, es importante señalar que, además de los adultos,
recientemente se han documentado casos clínicos de babesiosis en
becerros menores a nueve meses de edad, nacidos en áreas endémicas de la
enfermedad.
4 En el Centro Nacional de Investigación
Disciplinaria de Parasitología Veterinaria (CENID-PAVET) del Instituto
Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP)
del gobierno mexicano, se desarrolló una vacuna viva atenuada contra
Babesia bovis y B. bigemina, que protege al menos a 80% de los bovinos
vacunados contra cepas virulentas de Babesia
5-7 y a 70% de los animales bajo condiciones extremas de campo.
8
Por otro lado, se ha propuesto la utilización de la
bacteria ácido-láctica inmunoestimulante Lactobacillus casei como una
alternativa para el control de diferentes enfermedades parasitarias.9
En este contexto, se ha demostrado que L. casei, por sí misma, induce
una respuesta protectora contra Babesia microti en ratones cuando es
inoculada antes o el mismo día de la infección10 y contra
Babesia bovis y B. bigemina en ganado bovino, cuando la bacteria
ácido-láctica es administrada antes de la vacuna mixta contra babesiosis
bovina bajo condiciones controladas de laboratorio; sin embargo, L.
casei sola no indujo protección en el ganado vacuno contra el desafío
con Babesia.11
En el presente estudio se evaluó la efectividad de la
protección conferida por la administración simultánea de L. casei y la
vacuna mixta contra babesiosis bovina en bovinos que después de
vacunados fueron expuestos a la transmisión natural de Babesia bovis y
B. bigemina por la
garrapata
Rhipicephalus (Boophilus) microplus del ganado bajo condiciones
extremas en Paso del Toro, Veracruz, México, considerada como área
endémica de babesiosis bovina.
12
Material y métodos
Este estudio fue previamente revisado y aprobado de
acuerdo con las normas oficiales mexicanas NOM- 062-ZOO-1999,
NOM-033-ZOO-1995 y NOM-051- ZOO-1995 por miembros del subcomité de uso
ético y humanitario de animales del grupo técnico colegiado del
CENID-PAVET del INIFAP. Todos los bovinos usados en este experimento
fueron alojados, manejados y alimentados por personal entrenado de
acuerdo con las normas indicadas.
Vacuna contra babesiosis bovina
Se utilizó la vacuna bivalente atenuada contra
babesiosis bovina desarrollada por el INIFAP, que consiste en una mezcla
de la cepa atenuada BIS de Babesia bigemina y la clona BOR de B. bovis.8
Cepa bacteriana
La cepa ATCC7469 de Lactobacillus casei se cultivó
bajo condiciones aeróbicas en el medio MRS* a 37°C durante 18 h.
Posteriormente, los microorganismos fueron centrifugados a 5000 g por 10
min, y el precipitado se lavó varias veces con solución salina
amortiguadora estéril de fosfatos (PBS), más tarde, los organismos
murieron por calor en agua hirviendo durante 30 min y el número de
organismos en el cultivo se ajustó a 109 unidades formadoras de colonias
(CFU)/ml de PBS.13,14
Animales
Veinte bovinos cruza Holstein × Suizo, libres de
tuberculosis, brucelosis, babesiosis y anaplasmosis, con una edad
promedio de nueve meses, fueron distribuidos al azar en tres grupos:
no-vacunado: Testigo (n = 9), inmunizado con la vacuna mixta contra
babesiosis bovina: VAC (n = 5), e inoculado con VAC y Lactobacillus
casei: LC-VAC (n = 6).
Diseño experimental
Los animales se alojaron en un potrero en Coatepec,
Veracruz, ubicado a 1250 metros sobre el nivel del mar; coordenadas
geográficas: 19°25'0'' Norte, 96°47'0'' Oeste, donde recibieron los
siguientes tratamientos: los animales del grupo Testigo recibieron 3 ml
de solución salina estéril vía intramuscular (im) en el anca derecha;
los del grupo VAC fueron inmunizados vía im en el anca derecha con 1 ×
108 eritrocitos infectados con cada una de las especies de
Babesia del cultivo in vitro en un volumen de 3 ml, y los bovinos del
grupo LC-VAC fueron inoculados vía im en dos sitios diferentes: con VAC
(3 ml) en el anca derecha y con Lactobacillus casei (3 × 109
u.f.c.) en un volumen de 2 ml, en el anca izquierda. Luego se
mantuvieron en observación 24 días antes de transportarlos a Paso del
Toro, Veracruz (10 metros sobre el nivel del mar, coordenadas
geográficas: 19°2'0'' Norte, 96°7'0'' Oeste), un área endémica de
babesiosis bovina con una prevalencia estimada de babesiosis bovina
mayor a 90%,12 donde fueron expuestos en el campo a
garrapatas Rhipicephalus (Boophilus) microplus naturalmente infectadas
con Babesia bovis y Babesia bigemina. Esta situación se denominó
"condiciones extremas de campo", definidas como la suma de condiciones
de clima tropical, tiempo de verano, severa infestación de potreros con
garrapatas y área endémica de babesiosis bovina.
Registro de temperatura, volumen celular
aglomerado (hematocrito), eritrocitos parasitados con Babesia y niveles
de anticuerpos IgG anti-Babesia
En Paso del Toro, Veracruz, se registraron
diariamente los siguientes parámetros: temperatura rectal (°C) y volumen
celular aglomerado (PCV, como porcentaje). Se prepararon frotis
sanguíneos diariamente desde el día 14 después de la confrontación (dc),
teñidos con Giemsa para determinar por evaluación microscópica óptica
la presencia de Babesia spp y el porcentaje de eritrocitos parasitados
(PEP). Al mismo tiempo, se obtuvieron sueros de los bovinos los días 0 y
17 después de la vacunación (dv) y los días ocho y 20 dc para
determinar los niveles de anticuerpos IgG anti-Babesia bigemina y
anti-B-bovis por medio de la prueba de inmunofluorescencia indirecta
(IFI).15
Criterio para determinar babesiosis aguda
Un animal fue considerado con babesiosis aguda
(requiriendo tratamiento para evitar la muerte) cuando mostró los
siguientes parámetros: 1) No vacunado contra babesiosis bovina; 2) Más
de 25% de reducción del volumen celular aglomerado (VCA) con respecto al
valor basal; 3) Temperatura rectal mayor a 40°C durante dos días
seguidos; y 4) Presencia de Babesia spp en frotis sanguíneos teñidos con
Giemsa.5-8,11 Además, se tomó en cuenta la pobre condición física del animal (no medible).
Evaluación de signos clínicos de babesiosis y babesiosis aguda con riesgo de muerte
Similarmente, los porcentajes de signos clínicos de babesiosis y babesiosis aguda se determinaron de la siguiente manera:
• El porcentaje de signos clínicos de babesiosis =
número de animales con fiebre y presencia de parásitos en frotis teñidos
con Giemsa en el grupo/número total de animales en el grupo × 100.
• El porcentaje de babesiosis aguda con riesgo de
muerte = número de animales en el grupo que mostraron fiebre, reducción
de más de 25% del valor basal del VCA y presencia de parásitos en frotis
teñidos con Giemsa /número total de animales en el grupo × 100.
Es importante señalar que, en contraste con estudios
previos, los bovinos experimentales en el grupo Testigo se trataron
contra Babesia con 4,4'-(diamino) dibenzamide diaceturate* (Ganaseg)
cuando mostraron signos clínicos de babesiosis aguda después de la
confrontación, para evitar su muerte.6-8
Análisis estadístico
La significancia estadística de las diferencias se
determinó de las medias + desviación estándar (DE) por medio del
análisis de varianza (ANDEVA) con el
software Paquete de diseños experimentales.
16
Resultados
Temperatura
Desde el punto de vista clínico, los animales del
grupo VAC-LC mostraron un mejor desempeño en comparación con los bovinos
del grupo VAC, particularmente en el periodo de siete días, que abarcó
del día 14 al día 20 dc. En este periodo, el promedio de la temperatura
rectal (+ EE) fue significativamente más alto (P < 0.05) en los
animales del grupo VAC (40 + 0.08°C), en comparación con los bovinos del
grupo LC-VAC (39.5 + 0.1°C) (Figura 1).
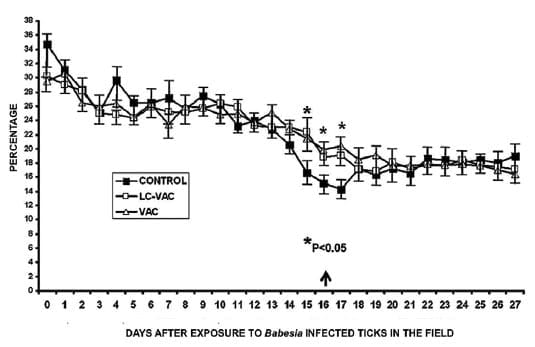
Volumen celular aglomerado (VCA)
En el día 0 no se observaron diferencias
significativas en este parámetro entre los grupos. Al día 15 dc el
porcentaje promedio de VCA en el grupo no-vacunado (Testigo) fue
significativamente más bajo (16.6%, P < 0.05), comparativamente con
los valores observados en los grupos VAC (21.5%) y LC-VAC (22.3%), y
representó una disminución de 52.4% con respecto al valor basal (día 0).
Los días 16 y 17, el VCA fue significativamente diferente en los grupos
vacunados, en comparación con el grupo Testigo (P < 0.05) (Figura
2).
Porcentaje de eritrocitos parasitados PEP
El día 10 dc se observó Babesia spp en los frotis
sanguíneos teñidos sólo en niveles inferiores a 0.01 PEP; luego los
valores se incrementaron, y los días 12 y 14 dc, se registró una
diferencia significativa (P < 0.01) en los valores promedio de PEP
observados en el grupo LC-VAC en comparación con los valores de los
grupos VAC y Testigo. Al día 12, los valores de PEP (promedio + EE)
fueron de 0.13% + 0.05 para el grupo LC-VAC; 0.53% + 0.04 para el grupo
VAC, y 0.43% + 0.12 para el grupo Testigo; mientras que los valores
observados el día 14 dc fueron de 0.10% + 0.05 para el grupo LC-VAC,
0.46% + 0.02 para el grupo VAC, y 0.45% + 0.12 para el grupo Testigo
(Figura 3).
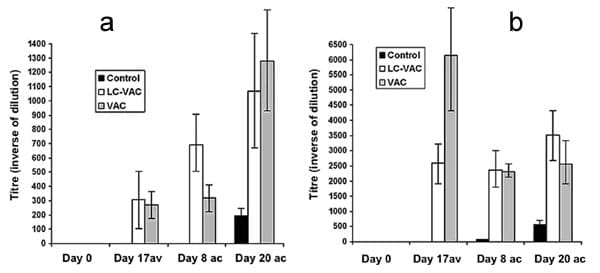
Niveles de anticuerpos IgG anti-Babesia
El promedio de los niveles de anticuerpos IgG
anti-Babesia en ambos grupos de animales vacunados siempre fue más alto
contra B. bovis que contra B. bigemina, después de la vacunación y
después de la confrontación. Al día ocho dc, el promedio de los niveles
de anticuerpos contra B. bigemina fue más alto en el grupo LCVAC que en
el grupo VAC (Figura 4a). Similarmente, el día 17 dv, el promedio de los
niveles de anticuerpos contra B. bovis fue más alto en el grupo VAC en
comparación con el grupo LC-VAC; mientras que el día 20 dc el promedio
de los niveles de anticuerpos contra B. bovis fue más alto en el grupo
LC-VAC que en el grupo VAC (Figura 4b); sin embargo, no se observaron
diferencias significativas. En el grupo Testigo el promedio de los
niveles de anticuerpos anti-Babesia dc fue muy bajo, comparado con los
promedios de los grupos LCVAC y VAC.
Figura 1. Temperatura promedio (°C)
en tres grupos de bovinos expuestos a Babesia spp transmitida por
Rhipicephalus (Boophilus) microplus en el campo. Cada punto representa
la media + E.E. de nueve (Testigo), seis (Lactobacillus casei + vacuna
mixta contra babesiosis: LC-VAC), y cinco bovinos (vacuna contra
babesiosis: VAC). Los asteriscos indican diferencias significativas (* P
< 0.01; ** P < 0.05) entre el grupo LC-VAC y los grupos Testigo o
VAC. Se administró tratamiento anti-Babesia a los animales Testigo el
día 16 (flecha).
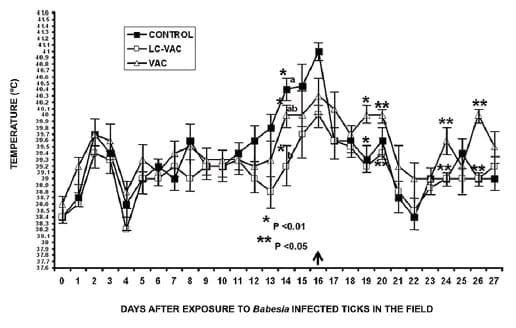
Figura 2. Porcentaje promedio del
volumen celular aglomerado (PCV) en tres grupos de bovinos expuestos a
Babesia spp transmitida por Rhipicephalus (Boophilus) microplus en el
campo. Cada punto representa la media + E.E. de nueve (Testigo), seis
(Lactobacillus casei + vacuna mixta contra babesiosis: LC-VAC) y cinco
bovinos (vacuna contra babesiosis: VAC). Los asteriscos indican
diferencias significativas (P < 0.05) entre el grupo LC-VAC y los
grupos Testigo o VAC. Se administró tratamiento anti-Babesia a los
animales Testigo el día 16 (flecha).
Signos clínicos de babesiosis y de babesiosis aguda con riesgo de muerte
Se observaron signos clínicos de babesiosis
(principalmente fiebre) el día 16 dc en los nueve animales (100%) del
grupo Testigo, en los seis bovinos (100%) del grupo VAC y en tres de
seis animales (50%) del grupo LC-VAC; mientras que se observaron signos
de babesiosis aguda con riesgo de muerte en los nueve bovinos (100%) del
grupo Testigo pero en ninguno de los animales (0%) de los grupos LC-VAC
y VAC. El mismo día, todos los animales del grupo Testigo mostraron
tanto B. bovis como B. bigemina en sus frotis sanguíneos teñidos con
Giemsa al ser examinados con el microscopio óptico. Al día 16 dc, tres
de nueve animales no vacunados estaban echados, mientras que todos los
bovinos de los grupos vacunados estaban de pie. Ese mismo día, todos los
animales en el grupo Testigo fueron tratados contra babesiosis con
4,4'-(diamino) dibenzamide diaceturate* (Ganaseg) durante dos días
consecutivos; a pesar del tratamiento, el día 26 dc un animal murió. Los
animales vacunados (grupos LCVAC y VAC) no recibieron tratamiento
contra babesiosis. El día 20 dc todos los animales fueron tratados
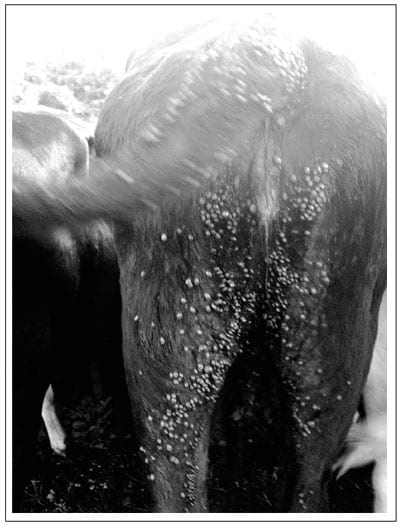
contra garrapatas (baño garrapaticida). Los bovinos mostraron cargas
promedio de 2000 garrapatas (oscilando entre 1500 a 3000 garrapatas por
animal). Un ejemplo de la carga de garrapatas se muestra en la Figura 5.
Figura 3. Porcentaje promedio de
eritrocitos parasitados por Babesia spp determinado por el examen al
microscopio de frotis sanguíneos teñidos con Giemsa, los días 12 y 14
después de la exposición de los tres grupos de bovinos a Babesia spp
transmitida por Rhipicephalus (Boophilus) microplus en el campo. Cada
punto representa la media + E.E. de nueve (Testigo), seis (Lactobacillus
casei + vacuna mixta contra babesiosis: LC-VAC), y cinco bovinos
(vacuna contra babesiosis: VAC). Los asteriscos indican diferencias
significativas (P < 0.01) entre el grupo LC-VAC y los grupos Testigo o
VAC.
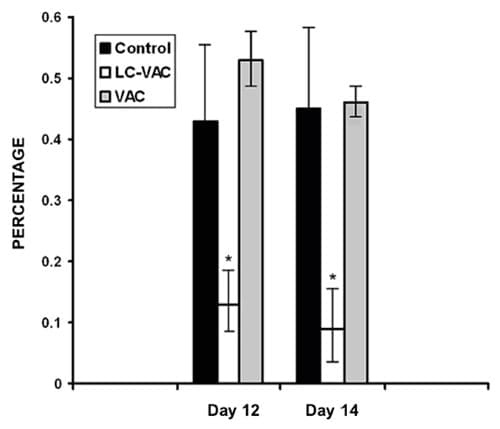
Figura 4. Promedio de los niveles de
anticuerpos IgG anti-Babesia, determinados por la prueba de
inmunofluorescencia indirecta (IFAT), los días 0 (valores basales) y 17
después de la vacunación (dv) y a los ocho y 20 después de la
confrontación (dc), de tres grupos de bovinos con Babesia spp
transmitida por Rhipicephalus (Boophilus) microplus en el campo. Cada
punto representa la media + E.E. de nueve (Testigo), seis (Lactobacillus
casei + vacuna mixta contra babesiosis: LC-VAC), y cinco bovinos
(vacuna contra babesiosis: VAC). a) IgG anti-Babesia bigemina; b) IgG
anti-Babesia bovis.
Discusión
Los resultados obtenidos demostraron la capacidad de
L. casei inoculado simultáneamente con la vacuna mixta contra babesiosis
bovina, para incrementar la eficiencia de dicha vacuna en animales (aun
en menores de nueve meses de edad) expuestos a la confrontación natural
con B. bovis y B. bigemina transmitidas por la garrapata Rhipicephalus
(Boophilus) microplus del ganado, y corroboraron los hallazgos previos
observados en bovinos vacunados y confrontados bajo condiciones
controladas con Babesia bovis y B. bigemina.11 En el estudio de Bautista et al.10
no se observó protección contra la confrontación con Babesia spp en
bovinos tratados únicamente con L. casei. Con base en esa observación,
en el presente estudio no se incluyó un grupo de bovinos tratados
solamente con L. casei. Similarmente, los resultados sugieren que los
bovinos del grupo VAC-LC están mejor protegidos contra la infección
natural con Babesia bovis y B. bigemina transmitidas por garrapatas, que
aquéllos que solamente recibieron la vacuna contra babesiosis. Se cree
que los incrementos en el promedio de temperatura observados los días 24
y 26 en el grupo VAC, se pudieron deber a una reinfección con Babesia
que luego fue controlada. Se sabe que L. casei estimula el sistema
inmunitario innato por medio de la activación de receptores tipo-Toll
(Toll-like receptors, TLRs) y la producción de citocinas tipo Th1.17,18
Los TLRs no solamente inducen respuestas inmunitarias innatas, sino que
también modulan la respuesta inmunitaria adaptativa celular y humoral
que da lugar a una mejor respuesta inmunitaria adquirida a un antígeno
en particular, como los antígenos de las vacunas.19,20 En
este sentido, se ha demostrado que una vacuna protectora de Leptospira
borgpetersenii muerta induce una potente inmunidad Th1 que comprende
respuestas mediadas por linfocitos T CD4 y T γδ T en el ganado vacuno.21,22
Figura 5.
Vista posterior de un bovino mostrando la carga de garrapatas el día 20
después de la introducción a un potrero infestado con Rhipicephalus
(Boophilus) microplus en La Posta Veracruz, México.
Los resultados observados en el presente estudio
sugieren que el uso de L. casei en vacunas contra babesiosis bovina
podría disminuir las severas respuestas a la vacunación que se observan
en el ganado vacunado.23
Es probable que la respuesta inmunitaria generada en
este estudio por L. casei y la vacuna bivalente contra babesiosis
consistió en una combinación de ambos brazos de la respuesta
inmunitaria, celular y humoral; sin embargo, los mecanismos celulares
deben ser investigados con más detalle en estudios futuros.
El bajo promedio de temperatura observado en el
periodo de siete días después de la exposición en el campo (días 14-20)
probablemente se debió a la menor presencia de parásitos en los animales
del grupo LC-VAC (39.5°C), en comparación con lo observado en los
bovinos del grupo VCA (40°C).
La caída en el VCA (particularmente durante los días
15, 16 y 17) se observó principalmente en los bovinos del grupo Testigo
por la infección de ambas especies de Babesia y la severa infestación
por garrapatas (Figura 5), mientras que la observada en los animales de
los grupos LC-VAC y VAC probablemente fue causada por la sola
infestación de garrapatas.
El PEP mostrado por los animales del grupo VAC,
similar al de los bovinos del grupo Testigo, sugiere que Babesia spp fue
finalmente controlada; sin embargo, los parásitos atenuados de la
vacuna sola generaron una respuesta protectora en los animales
vacunados, pero incapaz de evitar los signos clínicos temporales
causados por las cepas virulentas provenientes de la infección natural.
El bajo PEP observado en los bovinos del grupo LCVAC
en comparación con los valores mostrados en los animales de los grupos
Testigo y VAC probablemente se debió a la estimulación de una mejor
respuesta inmunitaria que controló la infección por Babesia con mayor
eficacia. En este contexto, es probable que la respuesta inmunitaria
innata fue activada por L. casei, como se ha demostrado en las
infecciones por B. microti. 10 Similarmente, se ha demostrado
que es importante la estimulación apropiada de la respuesta inmunitaria
innata en becerros, que involucra la inducción de citocinas Th1 y de
células tipo-NK en el bazo, para el control de B. bovis transmitida por
garrapatas R. (Boophilus) microplus.24
Aunque sólo se registró la producción de anticuerpos
IgG anti-Babesia en los animales de este estudio como manifestación de
la respuesta inmunitaria, es probable que la inmunidad celular haya
estado también involucrada en el control de B. bovis y B. bigemina en
los animales vacunados, como se ha sugerido anteriormente. 11,25,26
Se especula que las diferencias en los promedios de
los niveles de anticuerpos IgG anti-B. bovis y anti-B. bigemina en los
sueros de los animales vacunados en este sentido, posiblemente se
debieron a diferentes subclases de IgG estimuladas por los tratamientos;
es decir, el tratamiento simultáneo con L. casei y la vacuna contra
babesiosis estimuló diferentes subclases de IgG, a diferencia del
tratamiento con la vacuna sola. Sin embargo, se requiere llevar a cabo
más estudios en este sentido.
Es importante señalar que todos los animales en los
tres grupos fueron mantenidos en el potrero infestado con garrapatas
hasta por 16 días después de que se aplicó el tratamiento anti-Babesia
en los animales del grupo Testigo, periodo en el cual se observó una
disminución en el VCA en todos los animales. Respecto a ello, debe
tomarse en cuenta que el efecto directo de la infestación masiva con
garrapatas en los animales, que causa anemia y supresión del sistema
inmunitario de los bovinos,27,28 puede dar lugar a
reinfecciones con Babesia u otras infecciones (virales, bacterianas,
fúngicas) en animales débiles; por esta razón, es vital llevar a cabo
medidas de control de garrapatas, además de la vacunación contra Babesia
spp en zonas endémicas de babesiosis. Los resultados del presente
estudio concuerdan con investigaciones recientes en las cuales se ha
indicado que para mejorar las vacunas, los patrones moleculares
asociados con patógenos pueden ser utilizados para inducir respuestas
inmunitaria innatas, vía receptores tipo-Toll, que promueven la
inmunidad adaptativa.29 En este contexto, la observación
reciente de que casos clínicos de babesiosis se pueden presentar en
becerros menores de nueve meses de edad no-vacunados de una zona
endémica de babesiosis4 y los resultados del presente
trabajo, sugieren que el uso de L. casei puede proporcionar protección
adicional a la vacuna bivalente contra babesiosis bovina, tanto en
bovinos adultos como en becerros, contra la infección por Babesia en
explotaciones localizadas bajo condiciones tropicales donde la
babesiosis bovina generalmente es endémica.
Con base en los resultados obtenidos, se concluye
que: 1) la eficacia de la administración simultánea de L. casei y la
vacuna mixta mexicana contra babesiosis bovina fue mejor que la de la
vacuna sola, en términos de presentación de signos clínicos de la
enfermedad después de la exposición de animales vacunados a una
infestación masiva con garrapatas infectadas con B. bigemina y B. bovis
en un área endémica de babesiosis bovina; 2) la administración
simultánea de L. casei y la vacuna bivalente contra babesiosis bovina
genera una protección en los animales vacunados contra la infección
natural por Babesia, similar a la observada en bovinos tratados con L.
casei dos días antes de la administración de la vacuna bivalente contra
babesiosis bovina; 3) se requiere de más estudios para dilucidar los
mecanismos que ocurren en los animales vacunados con L. casei y la
vacuna bivalente contra babesiosis bovina; 4) los animales menores de
nueve meses de edad no vacunados contra babesiosis, desarrollan
babesiosis aguda cuando se exponen a una infestación masiva con
garrapatas infectadas con B. bovis y B. bigemina.
Agradecimientos
Este estudio fue financiado con fondos del proyecto
número 6216955P del Instituto Nacional de Investigaciones Forestales,
Agrícolas y Pecuarias (INIFAP).
Referencias
1. HUNFELD KP, HILDEBRANDT A, GRAY JS. Babesiosis: Recent insights into an ancient disease. Int J Parasitol 2008;38:1219-1237.
2. INEGI. Estados Unidos Mexicanos. Censo Agropecua
rio 2007, VIII Censo Agrícola, Ganadero y Forestal. Aguascalientes,
Ags., México: Instituto Nacional de Estadística y Geografía, 2009.
3. MOSQUEDA JJ. Vacunas contra hemoparásitos en
bovinos: Avances y perspectivas. En: BAUTISTA GARFIAS CR, FIGUEROA
MILLÁN JV, editores. Perspectivas de control de parásitos de importancia
veterinaria. CENID-Parasitología Veterinaria, INIFAP, Publicación
Técnica número 2. 2004;6-12.
4. OJEDA JJ, OROZCO L, FLORES R, ROJAS C, FIGUEROA
JV, ALVAREZ JA. Validation of an attenuated live vaccine against
babesiosis in native cattle in an endemic area. Transb Emerg Dis
2010;57:84-86.
5. CANTO GJ, FIGUEROA JV, ALVAREZ JA, VEGA CA.
Capacidad inmunoprotectora de una clona irradiada de Babesia bovis
derivada de cultivo in vitro. Tec Pecu Mex 1996;34:127-135.
6. FIGUEROA JV, CANTO GJ, ALVAREZ JA, LONA GR, RAMOS
JA, VEGA CA. Capacidad protectora en bovinos de una cepa de Babesia
bigemina derivada de cultivo in vitro. Tec Pecu Méx 1998;36:95-107.
7. ALVAREZ JA, RAMOS AJ, ROJAS E, MOSQUEDA JJ, VEGA
MCA, OLVERA A et al. Field challenge of cattle vaccinated with a
combined Babesia bovis and Babesia bigemina frozen immunogen. Ann N Y
Acad Sci 2004;1026:277-283.
8. CANTO GJ, ROJAS EE, ALVAREZ JA, RAMOS JA, MOSQUEDA
JJ, VEGA CA et al. Protection against bovine babesiosis with a mixed in
vitro culture derived B. bovis and B. bigemina vaccine under a field
challenge. Immunization in an endemic area. Tec Pecu Méx
2003;41:307-315.
9. BAUTISTA GARFIAS CR. Inmunoestimulación con
Lactobacillus casei como alternativa para el control de enfermedades
parasitarias. En: BAUTISTA GARFIAS CR, FIGUEROA MILLÁN JV, editores.
Perspectivas de control de parásitos de importancia veterinaria.
CENID-Parasitología Veterinaria, INIFAP, Publicación Técnica número 2.
2004:19-27.
10. BAUTISTA-GARFIAS CR, GOMEZ MB, AGUILAR BR, IXTA
O, MARTINEZ F, MOSQUEDA J. The treatment of mice with Lactobacillus
casei induces protection against Babesia microti infection. Parasitol
Res 2005; 97:472-427.
11. BAUTISTA CR, ALVAREZ JA, MOSQUEDA JJ, FALCON A,
RAMOS JA, ROJAS C et al. Enhancement of the Mexican bovine babesiosis
vaccine efficacy by using Lactobacillus casei. Ann N Y Acad Sci
2008;1149:126-130.
12. ÁLVAREZ MJA, CANTÓ AG. Epidemiología de la
babesiosis. En: QUIROZ ROMERO, H. Parasitología. México DF, Vol.
Conmemorativo de la Sociedad Mexicana de Parasitología SC, 1985:55-72.
13. BAUTISTA-GARFIAS CR, IXTA O, ORDUÑA M, MARTINEZ
F, AGUILAR B, CORTES A. Enhancement of resistance in mice treated with
Lactobacillus casei: Effect on Trichinella spiralis infection. Vet
Parasitol 1999;80:251-260.
14. BAUTISTA-GARFIAS C, IXTA-RODRIGUEZ O, MAR-
TINEZ-GOMEZ F, LOPEZ M, AGUILAR-FIGUEROA B. Effect of viable or dead
Lactobacillus casei organisms administered orally to mice on resistance
against Trichinella spiralis infection. Parasite 2001:8:226-228.
15. GUGLIELMONE AA, LUGARESIS CI, VOLPOGNI MM,
ANZIANA OS, VANZINI VR. Babesial antibody dynamics after cattle
immunization with live vaccines, measured with an indirect
immunofluorescence test. Vet Parasitol 1997;70:33-39.
16. OLIVAREZ SAENZ E. Paquete de diseños
experimentales, Marín (NL) México: Facultad de Agronomía de la
Universidad Autónoma de Nuevo León, México, 1994.
17. MALDONADO GALDEANO C, PERDIGON G. The probiotic
bacterium Lactobacillus casei induces activation of the gut mucosal
immune system through innate immunity. ClinVacc Immunol 2006;13:219-226.
18. VIZOSO PINTO MG, RODRIGUEZ GOMEZ M, SEIFERT S,
WATSI S, HOLZAPFEL WH, FRANZ CMAP. Lactobacilli stimulate the innate
immune response and modulate the TLR expression of HT29 intestinal
epithelial cells in vitro. Int J food Microbiol 2009;133:86-93.
19. BAUTISTA GARFIAS CR, MOSQUEDA GUALITO JJ. Role of
toll-like receptors in innate immunity and their implication in
veterinary medicine. Vet Mex 2005;36:453-468.
20. FERWERDA G, NETEA MG, JOOSTEN LA, VAN DER MEER
JWM, ROMANI L, KULLBERG BJ. The role of Toll-like receptors and C-type
lectins for vaccination against Candida albicans. Vaccine
2010;28:614-22.
21. NAIMAN BM, ALT D, BOLIN CA, ZUERNER R, BALDWIN C.
Protective killed Leptospira borgpetersenii vaccine induces potent Th1
immunity comprising responses by CD4 and γδ T lymphocytes. Infect Immun
2001;69:7550-7558
22. NAIMAN BM, BLUMERMAN S, ALT D, BOLIN CA, BROWN R
et al. Evaluation of type 1 immune response in naïve and vaccinated
animals following challenge with Leptospira borgpetersenii serovar
Hardjo: involvement of WC1+ γδ and CD4 T cells. Infect Immun
2002;70:6147-6157.
23. SHKAP V, LEIVOVITZ B, KRIGEL Y, HAMMERSCHLAG J,
MARCOVICS A, FISH L et al. Vaccination of older Bos taurus bulls against
bovine babesiosis. Vet Parasitol 2005;129:235-242.
24. GOFF WL, JOHNSON WC, HORN RH, BARRINGTON GM,
KNOWLES DP. The innate immune response in calves to Boophilus microplus
tick transmitted Babesia bovis involves type-1 cytokine induction and
NK-like cells in the spleen. Parasite Immunol 2003;25:185-188.
25. HOMER MJ, AGUILAR-DELFIN I, TELFORD III SR, KRAUSE PJ, PERSING DH. Babesiosis. Clin Microbiol Rev 2000;13:451-469.
26. AGUILAR-DELFIN I, WETTSTEIN PJ, PERSING DH.
Resistance to acute babesiosis is associated with interleukin-12- and
gamma interferon-mediated responses and requires macrophages and natural
killer cells. Infect Immunol 2003:71:2002-2008.
27. INOKUMA H, KERLIN RL, KEMP DH, WILLADSEN P.
Effects of cattle tick (Boophilus microplus) infestation on the bovine
immune system. Vet Parasitol 1993;47:107-118.
28. JONSSON NN. The productivity effects of cattle
tick (Boophilus microplus) infestation on cattle, with particular
reference to Bos indicus cattle and their crosses. Vet Parasitol
2006;137:1-10.
29. BACHMANN MF, JENNINGS CT. Vaccine delivery: a
matter of size, geometry, kinetics and molecular patterns. Nat Rev
Immunol 2010;10:787-796.